NOTES FROM THE FIELD / NOTES DU TERRAIN
Fundamental Concepts Bridging Education and the Brain
STEVE MASSON & LORIE-MARLÈNE BRAULT FOISY
Université du Québec à Montréal
ABSTRACT. Although a number of papers have already discussed the relevance of brain research for education, the fundamental concepts and discoveries connecting education and the brain have not been systematically reviewed yet. In this paper, four of these concepts are presented and evidence concerning each one is reviewed. First, the concept of neuroplasticity is proposed as a sine qua non for linking education and the brain. Then, the concepts of neuronal recycling and inhibition are presented as two fundamental mechanisms of school learning that emphasize the importance of knowing the initial brain structure of learners and, finally, the concept of attention is discussed as a central concept for linking teaching and the brain.
COncepts fondamentaux permettant d’établir des ponts entre l’éducation et le cerveau
RÉSUMÉ. Bien qu’un certain nombre d’articles aient déjà discuté de la pertinence des recherches sur le cerveau pour le domaine de l’éducation, les découvertes et les concepts fondamentaux reliant l’éducation et le cerveau n’ont jamais fait l’objet d’une analyse systématique. Dans cet article, quatre de ces concepts sont présentés et, pour chacun de ces concepts, une brève synthèse de la littérature est proposée. Premièrement, le concept de neuroplasticité est proposé comme une condition sine qua non pour établir des liens entre l’éducation et le cerveau. Ensuite, les concepts de recyclage neuronal et d’inhibition sont présentés comme deux mécanismes fondamentaux liés aux apprentissages scolaires qui mettent l’accent sur l’importance de connaître la structure initiale du cerveau des apprenants et, enfin, le concept d’attention est présenté comme un concept central pour établir des liens entre l’enseignement et le cerveau.
The number of studies on the human brain has increased exponentially over the last 20 years. As research in neuroscience progressed, the educational community has become intrigued by how this new knowledge might contribute to a better understanding of school learning, and even potentially contribute to the improvement of teaching (Organisation for Economic Co-operation and Development [OECD], 2007). This growing interest in the role of neuroscience in education has led to the creation of a field of research called neuroeducation, educational neuroscience, “mind, brain, and education,” or simply “neuroscience and education.” This field aims to establish connections between education and the brain (Fisher, Daniel, & Immordino-Yang, 2007; Masson, 2012), with the ultimate goal of improving certain educational issues.
In general, researchers in neuroscience and education agree that understanding how the brain works might provide some interesting insights into education (Ansari, Smedt, & Grabner, 2011; Fisher et al., 2007; The Royal Society, 2011), and even contribute to the advancement of evidence-based education (Pasquinelli, 2011). However, they also agree that connecting education and the brain is not a simple task because there is a gap between the brain and the classroom that cannot be bridged easily (Bruer, 2006; Willingham, 2009).
Some papers have provided pertinent guidelines and strategies for research aiming to link the brain and education (Varma, McCandliss, & Schwartz, 2008; Willingham & Lloyd, 2007). However, the fundamental concepts supporting the claim that studying education with neuroscientific tools such as brain imaging is not only possible but also potentially useful for education have not been systematically reviewed yet. For this reason, this paper proposes an answer to the question, what are the fundamental concepts and relationships connecting education and the brain? by discussing four core concepts that contribute to bridging the gap between neuroscience and education: neuroplasticity, neuronal recycling, inhibition and attention. These concepts and their relationships are summarized in Figure 1.
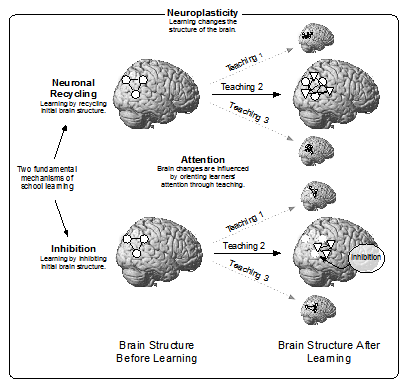
FIGURE 1. Fundamental concepts and relationships connecting education and the brain
Neuroplasticity refers to the capacity of the brain to change its structure (its neuronal connections, more precisely) through learning. School learning is influenced by learners’ initial brain structures and by at least two learning mechanisms: (a) neuronal recycling (in which the function of evolutionarily older brain circuits is modified by new cultural inventions such as reading) and (b) inhibition (in which neuronal networks — that are hard to change and that lead to inappropriate responses — are deactivated by frontal brain regions related to inhibition). Attention is an essential concept for linking teaching and the brain; brain changes during learning are influenced by the way learners’ attention is directed through teaching.
NEUROPLASTICITY
Neuroplasticity is one of the most important discoveries in recent neuroscience. Although different meanings of this concept exist (Berlucchi & Buchtel, 2009), in this paper, it refers to the ability of the brain to change its neural connections through learning. Neuroplasticity is the sine qua non for linking education and the brain. Indeed, if learning did not change the brain, it would be impossible to study how education affects the brain because no change could be observed with brain imaging or other neuroscientific techniques.
Fortunately, for neuroeducational research, there is strong evidence that learning changes the structure of the brain. For example, Maguire et al. (2000) have studied the brains of London taxi drivers who have extensive navigation experience. They found that the hippocampal volume of taxi drivers was correlated to the amount of time spent as a taxi driver. In another study, Draganski et al. (2004) observed that people who learned to juggle over a three-month period showed an increased grey matter density in brain regions related to visual motion. More recently, Kwok et al. (2011) showed that adults who learned new colour names increased their grey matter density in visual areas involved in colour processing, even after only a short two-hour training session.
Neuroplasticity is not only a necessary condition for linking education and the brain, it also has some interesting implications for education, as Geake and Cooper (2003) pointed out. These implications result largely from the work of Hebb (1949) who was one of the first to propose that underlying modifications of neural connections in the brain cause changes in behaviour. He stated that:
When an axon of cell A is near enough to excite cell B and repeatedly or persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A’s efficiency, as one of the cells firing B, is increased. (p. 62)
Hebb’s rule led to the well-known principle that neurons that fire together, wire together.
According to Geake and Cooper (2003), even if not all types of learning respect this principle, Hebb’s model provides interesting information for school learning. First, it explains why school learning takes time and practice: neurons need to fire together repeatedly or persistently in order to increase their connections with each other. Second, it also explains why students forget: when neurons do not fire together for some time, the strength of their connections decreases progressively. Third, Hebb’s model explains why some mistakes, systematically made by students, can be hard to change: they might be induced by neuronal networks that are strongly established in the brain (probably because they are useful and reinforced in other contexts) and that cannot be changed easily. For example, science teachers are often confronted with students’ initial conceptions about various natural phenomena. These initial conceptions can be really resistant to change, making some science concepts particularly difficult to learn for students. Although these initial conceptions are not in line with the accepted scientific models, they might be sufficiently effective (and even useful in daily life) to make them deeply entrenched (Limon, 2001). Students often think, for instance, that it is warmer during summer because the Earth is closer to the Sun (which is scientifically incorrect). This mistake could be due to the fact that this idea is often reinforced in everyday life when observing that an object closer to a heat source becomes warmer (Salierno, Edelson, & Sherin, 2005).
NEURONAL RECYCLING
Although the concept of neuroplasticity establishes a strong and fundamental relationship between education and the brain, the implications of brain research for education would be limited if the brain was totally plastic and able to easily modify any of its areas to acquire any new ability. Indeed, why would knowing about the brain help educators to improve their teaching if students’ brains were so flexible that they could efficiently learn anything through any teaching method (Dehaene, 2008)?
Recent advances in neuroscience have provided growing evidence that the brain is not completely plastic and that some brain areas are more predisposed than others to acquiring new abilities (Dehaene, 2005; Dehaene & Cohen, 2007). For example, learning to read does not modify various parts of the brain, depending on each individual; it modifies only one. More precisely, studies show that reading letters and words is associated with systematic activation in the left occipito-temporal cortex, also called the visual word form area (VWFA) (Dehaene, Le Clec’H, Poline, Le Bihan, & Cohen, 2002; Gaillard, Balsamo, Ibrahim, Sachs, & Xu, 2003; Marinkovic et al., 2003). In fact, this region seems universally related to reading across different languages, even in Chinese (Nakamura et al., 2012). Studies also show that activation in this region is correlated with reading performance (Beaulieu et al., 2005), age (Shaywitz et al., 2007) and education (Dehaene et al., 2010). Although the left occipito-temporal region is not the only area that can change to develop people’s ability to read (Seghier, 2012), it seems easier and more natural for the brain to establish in this region the neural connections which allow reading.
Why does the brain show a preference for the left occipito-temporal cortex? Dehaene argues that learning is influenced by previous neuronal organization within the brain, which is itself influenced by genetic and epigenetic factors (Dehaene & Cohen, 2007). There are at least two reasons why the left occipito-temporal cortex is predisposed to reading. First, since reading involves visual processing and the optic nerve is connected to the occipital cortex, it seems plausible that brain areas devoted to reading preferentially invade regions near the primary visual cortex. However, the VWFA cannot be established directly beside the visual cortex because the ability to read requires neurons that activate regardless of the size and style of letters, which requires distance from the primary sensory brain areas. Second, when children learn to read, most of them have already developed brain areas related to speech language in the left hemisphere. Consequently, it seems plausible to think that the VWFA must be situated in the left hemisphere to allow an easier connection between visual and language brain circuits.
The concept of “neuronal recycling” proposed by Dehaene (2005) describes the process by which evolutionarily older brain circuits are invaded by new cultural inventions such as reading. It can be seen as a second fundamental concept in neuroeducation. According to this perspective, students must modify or recycle brain circuits already in place in the brain in order to learn to read and count. Because some brain circuits can be recycled to develop certain abilities more easily than others, neuroscience can provide useful information for educators by identifying and understanding the biological constraints that the brain organization imposes on how certain school abilities can be learned.
INHIBITION
As discussed in the previous section, initial brain structure and neuronal networks affect learning. Sometimes, this initial structure helps in the learning of new abilities. For instance, when learning to read letters and words, the brain uses neuronal networks that are able to identify objects and optimizes (or recycles) them in order to identify a particular class of objects — in this case, letters and words. However, sometimes the initial brain structure can be an obstacle to learning because it can bias students’ reasoning and lead to incorrect responses that can be hard to modify. In such cases, learning requires the inhibition of the spontaneous activation of some neuronal networks that are inappropriate for the task. Inhibition is a third fundamental concept in neuroeducation. It can also be viewed as the second fundamental mechanism of school learning (the first being neuronal recycling).
Over the last 30 years, research in science and mathematics education has shown that students often have misconceptions about mathematics and how nature works (Confrey, 1990). Most of the time, these conceptions exist even before students have received formal instruction on the subject, and they can be particularly hard to modify (diSessa, 2006). Recent studies using neuroimaging have revealed that overcoming these misconceptions or intuitions requires the activation of brain areas related to inhibition such as the ventrolateral prefrontal cortex (VLPC), the dorsolateral prefrontal cortex (DLPC) and the anterior cingulate cortex (ACC).
In mathematics, Houdé et al. (2011) showed that young children (5–6 years old) have difficulty understanding that two rows that contain the same number of objects, but that have different lengths because of differential spacing between the objects (e.g., oooooo and o o o o o o), indeed contain the same number of objects. At the age of 9 to 10 years old, this spontaneous and wrong answer is usually overcome: children respond that the two rows contain the same number of objects. Among the brain areas that are more activated in successful 9 to 10-year-olds compared to unsuccessful 5 to 6-year-olds, there are brain areas related to number sense (e.g., intraparietal sulcus), but also to inhibition, probably because the brain must inhibit the biased tendency to consider that longer rows have more objects. At this time, we ignore whether this need of inhibition persists during adolescence and adulthood. However, the same team of researchers (Houdé et al., 2000) also found that adults who were trained to overcome a perceptual bias leading to wrong answers in a deductive logic task showed more activation after rather than before training in frontal lobe areas involved in inhibition. Other researchers (Stavy & Babai, 2010) found that the activation of brain areas related to inhibition is also correlated to the capacity to correctly answer counter-intuitive questions about the perimeter of figures.
Research in science education has revealed similar results. A recent study by Masson, Potvin, Riopel, and Brault Foisy (2014) shows that experts in physics who overcome the common misconception “one wire is sufficient to light a bulb” have stronger activation in brain areas usually involved in inhibition (such as the ventrolateral prefrontal cortex, the dorsolateral prefrontal cortex and the anterior cingulate cortex) than novices in physics when they answer questions related to this misconception. Another study reveals that the activation of inhibition’s brain areas is also observed during a task involving a common misconception about mechanics (“heavier objects fall faster”) (Brault Foisy, Potvin, Riopel, & Masson, 2014). These results, combined with other behavioural data (Shtulman & Valcarcel, 2012), suggest that experts in science still have misconceptions in their brains that have not been erased or replaced during science education. Instead, scientific learning seems to develop the ability to inhibit neuronal networks associated with misconceptions.
ATTENTION
The concepts of neuroplasticity, neuronal recycling, and inhibition provide solid foundations for linking education and the brain. However, if neuroeducation aims to improve education through research on the brain, knowing about how students’ brains change with learning is not sufficient. A relationship must be established between teaching practices and what happens inside students’ brains during learning. As proposed by McCandliss (2011), we think that attention is a central concept in the establishment of a bridge between teaching and the learning brain, not only because attention is a shared concept between education and neuroscience, but also because research shows that when a teacher directs a student’s attention during learning, it influences how the brain works to accomplish a task and learn new abilities.
A study by Yoncheva, Blau, Maurer, and McCandliss (2010) provides a clear example of how teaching can influence learners’ brain activity. In their study, two groups of adults were asked to learn a new writing system created solely for the purpose of the research. This system consisted of several symbols (words) formed of three letters (graphemes), each of which was connected to a specific sound (phoneme). The letters embedded in the symbols were evident only when instructions draw attention to them. In the first group (whole-word group), people were only taught that each symbol corresponded to a particular word, and so they learned to read the list of words by directing their attention to the whole symbol. In the second group (grapheme-phoneme group), people had to learn to read the same list of words, but this time, the teacher directed the learners’ attention toward the sub-parts of the symbol by saying that the symbol was composed of three parts and that the lines at the bottom, middle and top corresponded respectively to the first, second and third phonemes of the word. The design of the research allows to isolate the influence of attentional focus during teaching. Figure 2 presents the instructions given to each group for one symbol.
Although there was only one subtle change in how reading was taught, the results revealed important differences in brain activity between the whole-word group and the grapheme-phoneme group. Indeed, the whole-word group showed more brain activity in the right hemisphere 150 ms after the presentation of a symbol, whereas the grapheme-phoneme group showed more activity in the left hemisphere, near the occipito-temporal region. Since this brain area is strongly correlated with expertise in reading (Beaulieu et al., 2005), this result suggests that the grapheme-phoneme approach might help learners to build an efficient reading neuronal network, while the whole-word approach might not.
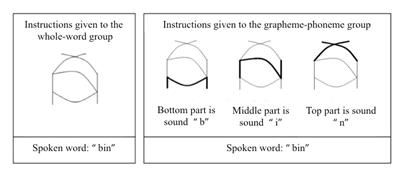

FIGURE 2. Instructions given to each group of participants for the symbol “bin”
Other studies provide evidence that teaching methods influence learners’ brains. For example, teaching deductive logic with or without warnings about a potential perceptual bias produces different brain activity (Houdé et al., 2001). In mathematics, teaching new arithmetic operations by strategies or by drill also produces differential modifications in brain activation (Delazer et al., 2005), as do orthographic or morphological spelling treatments in children with dyslexia (Richards et al., 2006). It is important to note that differential modifications of brain activation do not necessarily imply that different teaching methods produce different structural changes in learners’ brains. However, since “neurons that fire together, wire together,” it appears plausible that differential brain activation produces differential structural modifications in the brain. But, to our knowledge, no study has shown yet that two types of teaching produce different changes in brain structure. Further research is therefore needed to consolidate the hypothesis that teaching can influence how the brain changes through learning.
CONCLUSION
This papers aims to identify and discuss the fundamental concepts and discoveries connecting education and the brain. Although neuroplasticity is perhaps the most fundamental of these concepts, neuronal recycling, inhibition and attention are also quite important — the first two because they reveal the importance of understanding the biological constraints that the initial brain structure imposes on learning, and the last one because it provides a connection between what teachers do and what happens inside learners’ brains.
To strengthen the relationships between education and the brain, further work is needed in at least two domains. First, to our knowledge, although studies have shown that different teaching methods induce differential changes in brain activations, there is no direct evidence that these changes produce modifications to the brain’s structure. Second, there are many studies about how the brain develops cultural abilities such as reading and counting, but only a few studies about learning content that is difficult and counter-intuitive for students, such as certain scientific concepts. Studying how the brain overcomes persistent errors, and how inhibition training can help to overcome these difficulties, might have important consequences for learning and teaching.
REFERENCES
Ansari, D., Smedt, B., & Grabner, R. H. (2011). Neuroeducation – A critical overview of an emerging field. Neuroethics. doi: 10.1007/s12152-011-9119-3
Beaulieu, C., Plewes, C., Paulson, L. A., Roy, D., Snook, L., Concha, L., & Phillips, L. (2005). Imaging brain connectivity in children with diverse reading ability. NeuroImage, 25(4), 1266-1271.
Berlucchi, G., & Buchtel, H. A. (2009). Neuronal plasticity: historical roots and evolution of meaning. Exp Brain Res, 192(3), 307-319. doi: 10.1007/s00221-008-1611-6
Brault Foisy, L.-M., Potvin, P., Riopel, M., & Masson, S. (2014). Do Experts in Mechanics Still Inhibit Common Physics Misconceptions? Manuscript submitted for publication.
Bruer, J. T. (2006). On the implications of neuroscience research for science teaching and learning: Are there any? A skeptical theme and variations: The primacy of psychology in science learning. CBE-Life Science Education, 5, 104-110.
Confrey, J. (1990). A review of the research on student conceptions in mathematics, science, and programming. In C. B. Cazden (Ed.), Review of Research in Education (Vol. 16, pp. 3-56). Washington, NW: American Educational Research Association.
Dehaene, S. (2005). Evolution of human cortical circuits for reading and arithmetic: The “neuronal recycling” hypothesis. In S. Dehaene, J. R. Duhamel, M. Hauser, & G. Rizzolatti (Eds.), From monkey brain to human brain (pp. 133-157). Cambridge, MA: MIT Press
Dehaene, S. (2008). Cerebral constraints in reading and arithmetic: Education as a “neuronal recycling” process. In A. M. Battro, K. W. Fischer, & P. Léna (Eds.), The educated brain: Essays in neuroeducation (pp. 232-247). Cambridge, UK: Cambridge University Press
Dehaene, S., & Cohen, L. (2007). Cultural recycling of cortical maps. Neuron, 56(2), 384-398. doi: 10.1016/j.neuron.2007.10.004
Dehaene, S., Le Clec’H, G., Poline, J.-B., Le Bihan, D., & Cohen, L. (2002). The visual word form area: a prelexical representation of visual words in the fusiform gyrus. NeuroReport, 13(3), 321-325.
Dehaene, S., Pegado, F., Braga, L. W., Ventura, P., Filho, G. N., Jobert, A.,... Cohen, L. (2010). How learning to read changes the cortical networks for vision and language. Science, 330(6009), 1359-1364. doi: 10.1126/science.1194140
Delazer, M., Ischebeck, A., Domahs, F., Zamarian, L., Koppelstaetter, F., Siedentopf, C. M.,... Felber, S. (2005). Learning by strategies and learning by drill — evidence from an fMRI study. NeuroImage, 25(3), 838-849.
diSessa, A. A. (2006). A history of conceptual change research: Threads and fault lines. In R. K. Sawyer (Ed.), Cambridge handbook of the learning sciences (pp. 265-281). Cambridge, UK: Cambridge University Press
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., & May, A. (2004). Neuroplasticity: Changes in grey matter induced by training. Nature, 427(6972), 311-312. doi: 10.1038/427311a
Fischer, K. W., Daniel, D. B., Immordino-Yang, M. H., Stern, E., Battro, A., & Koizumi, H. (2007). Why Mind, Brain, and Education? Why Now? Mind, Brain, and Education, 1(1), 1-2. doi: 10.1111/j.1751-228X.2007.00006.x
Gaillard, W. D., Balsamo, L. M., Ibrahim, Z., Sachs, B. C., & Xu, B. (2003). fMRI identifies regional specialization of neural networks for reading in young children. Neurology, 60(1), 94-100.
Geake, J., & Cooper, P. (2003). Cognitive neuroscience: Implications for education. Westminster Studies in Education, 26(1), 7-20.
Hebb, D. O. (1949). The organization of behavior. New York, NY: Wiley.
Houdé, O., Pineau, A., Leroux, G., Poirel, N., Perchey, G., Lanoe, C.,... Mazoyer, B. (2011). Functional magnetic resonance imaging study of Piaget’s conservation-of-number task in preschool and school-age children: A neo-Piagetian approach. Journal of experimental child psychology, 110(3), 332-346. doi: 10.1016/j.jecp.2011.04.008
Houdé, O., Zago, L., Crivello, F., Moutier, S., Pineau, A., Mazoyer, B., & Tzourio-Mazoyer, N. (2001). Access to deductive logic depends on a right ventromedial prefrontal area devoted to emotion and feeling: Evidence from a training paradigm. NeuroImage, 14, 1486-1492.
Houdé, O., Zago, L., Mellet, E., Moutier, S., Pineau, A., Mazoyer, B., & Tzourio-Mazoyer, N. (2000). Shifting from the perceptual brain to the logical brain: The neural impact of cognitive inhibition training. Journal of Cognitive Neuroscience, 12(5), 721-728.
Kwok, V., Niu, Z., Kay, P., Zhou, K., Mo, L., Jin, Z.,... Tan, L. H. (2011). Learning new color names produces rapid increase in gray matter in the intact adult human cortex. Proceedings of the National Academy of Sciences of the United States of America, 108(16), 6686-6688. doi: 10.1073/pnas.1103217108
Limon, M. (2001). On the cognitive conflict as an instructional strategy for conceptual change: A critical appraisal. Learning and Instruction, 11, 357-380.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S., & Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences, 97(8), 4398-4403. doi: 10.1073/pnas.070039597
Marinkovic, K., Dhond, R. P., Dale, A. M., Glessner, M., Carr, V., & Halgren, E. (2003). Spatiotemporal dynamics of modality-specific and supramodal word processing. Neuron, 38(3), 487-497.
Masson, S. (2012). Neuroeducation: Understanding the brain to improve teaching. Neuroeducation, 1(1), 1-2.
Masson, S., Potvin, P., Riopel, M., & Brault Foisy, L.-M. (2014). Do experts in electricity still have a misconception in their brains that must be inhibited? Mind, Brain, and Education, 8(1), 44-55. doi: 10.1111/mbe.12043
McCandliss, B. D. (2011, June). Selective attention as a bridge between education and neuroscience: How differences in instruction change the brain circuitry involved in learning to read. Paper presented at the Third Conference of the International Mind, Brain, and Education Society (IMBES), San Diego, CA, USA.
Nakamura, K., Kuo, W.-J., Pegado, F., Cohen, L., Tzeng, O. J. L., & Dehaene, S. (2012). Universal brain systems for recognizing word shapes and handwriting gestures during reading. Proceedings of the National Academy of Sciences, 109(50), 20762–20767. doi: 10.1073/pnas.1217749109
Organisation for Economic Co-operation and Development (2007). Understanding the brain: The birth of a learning science. Paris: Author.
Pasquinelli, E. (2011). Knowledge- and evidence-based education: Reasons, trends, and contents. Mind, Brain, and Education, 5(4), 186-195. doi: 10.1111/j.1751-228X.2011.01128.x
Richards, T. L., Aylward, E. H., Berninger, V. W., Field, K. M., Grimme, A. C., Richards, A. L., & Nagy, W. (2006). Individual fMRI activation in orthographic mapping and morpheme mapping after orthographic or morphological spelling treatment in child dyslexics. Journal of Neurolinguistics, 19(1), 56-86.
Salierno, C., Edelson, D., & Sherin, B. (2005). The development of student conceptions of the earth-sun relationship in an inquiry-based curriculum. Journal of Geoscience Education, 53(4), 422.
Seghier, M. L., Neufeld, N. H., Zeidman, P., Leff, A. P., Mechelli, A., Nagendran, A.,... Price, C. J. (2012). Reading without the left ventral occipito-temporal cortex. Neuropsychologia, 50(14), 3621-3635. doi: 10.1016/j.neuropsychologia.2012.09.030
Shaywitz, B. A., Skudlarski, P., Holahan, J. M., Marchione, K. E., Constable, R. T., Fulbright, R. K.,... Shaywitz, S. E. (2007). Age-related changes in reading systems of dyslexic children. Annals of Neurology, 61(4), 363-370. doi: 10.1002/ana.21093
Shtulman, A., & Valcarcel, J. (2012). Scientific knowledge suppresses but does not supplant earlier intuitions. Cognition, 124(2), 209-215. doi: 10.1016/j.cognition.2012.04.005
Stavy, R., & Babai, R. (2010). Overcoming intuitive interference in mathematics: Insights from behavioral, brain imaging, and intervention studies. ZDM Mathematics Education, 42(6), 621-633. doi: 10.1007/s11858-010-0251-z
The Royal Society. (2011). Neuroscience: Implications for education and lifelong learning. London, UK: The Royal Society.
Varma, S., McCandliss, B. D., & Schwartz, D. L. (2008). Scientific and pragmatic challenges for bridging education and neuroscience. Educational Researcher, 37(3), 140-152. doi: 10.3102/0013189x08317687
Willingham, D. T. (2009). Three problems in the marriage of neuroscience and education. Cortex, 45(4), 544-545. doi: 10.1016/j.cortex.2008.05.009
Willingham, D. T., & Lloyd, J. W. (2007). How educational theories can use neuroscientific data. Mind, Brain, and Education, 1(3), 140-149. doi: 10.1111/j.1751-228X.2007.00014.x
Yoncheva, Y. N., Blau, V. C., Maurer, U., & McCandliss, B. D. (2010). Attentional focus during learning impacts N170 ERP responses to an artificial script. Developmental neuropsychology, 35(4), 423 - 445. doi: 10.1080/87565641.2010.480918